Virus dell’Influenza: Ho Spiato Come Riprogramma le Tue Cellule in Diretta!
Ciao a tutti! Oggi voglio portarvi con me in un viaggio affascinante nel mondo microscopico, anzi, nanoscopico, per scoprire cosa succede esattamente quando un singolo, minuscolo virus dell’influenza A incontra una delle nostre cellule. È un momento cruciale, l’inizio dell’infezione, ma è anche incredibilmente fugace e difficile da osservare. Immaginate di dover studiare una stretta di mano che dura una frazione di secondo, avvenuta in uno spazio minuscolo!
La Sfida: Catturare l’Attimo Fuggente
Quando un virus dell’influenza A (che chiameremo IAV per comodità) ci infetta, la prima cosa che fa è aggrapparsi alla superficie delle nostre cellule, in particolare a quelle del nostro sistema respiratorio. Lo fa usando delle proteine sulla sua superficie, chiamate emoagglutinine (HA), che si legano a specifiche molecole zuccherine (gli acidi sialici) presenti sulla membrana delle nostre cellule. Questo legame iniziale, però, è piuttosto debole. Come fa allora il virus a non essere spazzato via? Abbiamo scoperto di recente che i virus sono furbi: non si legano a un solo recettore, ma a gruppi di recettori, come se usassero più mani per tenersi saldi.
Ma cosa succede subito dopo? Il virus deve entrare nella cellula per potersi replicare. Questo processo, chiamato endocitosi, è spesso mediato da una proteina chiamata clatrina (si parla di endocitosi mediata da clatrina o CME). Il virus, in pratica, “convince” la cellula a inglobarlo. Ma come fa a inviare questo segnale? I recettori a cui si lega inizialmente non hanno capacità di segnalazione diretta. Si è ipotizzato che altre proteine cellulari, come dei recettori speciali chiamati “recettori tirosin-chinasi” (ad esempio l’EGFR, recettore del fattore di crescita epidermico), vengano coinvolte. Studi precedenti avevano suggerito che l’EGFR potesse avere un ruolo nell’ingresso dell’IAV, ma vedere questo processo in azione, al livello del singolo virus e in tempo reale, era una sfida enorme. Gli eventi chiave avvengono in pochi minuti e su scale così piccole (nanometri!) che le tecniche tradizionali faticano a coglierne i dettagli spazio-temporali.
La Nostra Idea: “Incollare” il Virus per Studiarlo Meglio
Per superare questi limiti, abbiamo pensato: e se potessimo “fermare il tempo”, o almeno rallentarlo un po’? Abbiamo sviluppato un metodo per immobilizzare chimicamente i virus influenzali, senza modificarli troppo, su dei vetrini da microscopio. In pratica, abbiamo creato una superficie “appiccicosa” solo per i virus. Poi, su questi virus “incollati”, abbiamo fatto crescere delle cellule epiteliali umane vive (le cellule A549, una linea cellulare comune per studiare le infezioni polmonari).
In questo modo, l’interfaccia tra il virus e la membrana cellulare rimane stabile per un tempo molto più lungo, permettendoci di usare tecniche di microscopia avanzate, come la microscopia a super-risoluzione a singola molecola (SMLM), per osservare cosa succede in dettaglio. È un po’ come mettere in pausa una scena d’azione per analizzarne ogni fotogramma! Per assicurarci che anche le cellule si attaccassero bene al vetrino, abbiamo aggiunto una piccola molecola (chiamata cRGD) che aiuta le cellule ad aderire, un po’ come un nastro biadesivo molecolare. Abbiamo verificato con la microscopia confocale che le cellule crescessero effettivamente sopra i virus immobilizzati, creando l’assetto sperimentale perfetto.

Spiare le Interazioni Virus-Recettore: L’EGFR Sotto la Lente
Armati di questa nuova piattaforma, ci siamo concentrati sull’interazione tra l’IAV e il recettore EGFR. Usando cellule che esprimevano EGFR marcato con una proteina fluorescente (EGFR-GFP), abbiamo osservato al microscopio TIRF (una tecnica che illumina solo un sottilissimo strato vicino al vetrino, perfetto per vedere la membrana cellulare) che l’EGFR si accumulava proprio dove erano immobilizzati i virus. E questa accumulazione dipendeva dal legame iniziale del virus ai recettori con acido sialico: se trattavamo le cellule con un enzima (sialidasi) che rimuove questi zuccheri, l’accumulo di EGFR spariva. Questo confermava un legame specifico!
Ma volevamo andare ancora più a fondo. Abbiamo usato una tecnica di super-risoluzione chiamata PALM (Photoactivated Localization Microscopy), combinata con il tracciamento di singole particelle (sptPALM). In pratica, abbiamo “acceso” e localizzato singole molecole di EGFR (marcate con un’altra proteina fluorescente, mEos3.2) una alla volta, ricostruendo i loro movimenti con precisione nanometrica. È come seguire ogni singolo ballerino su una pista affollata!
Cosa abbiamo scoperto? Analizzando migliaia di traiettorie di EGFR, abbiamo visto che, in generale, la presenza dei virus non alterava la mobilità media dei recettori su tutta la cellula. Ma, guardando specificamente nelle immediate vicinanze dei virus (in un quadratino immaginario di 800×800 nanometri attorno a ogni virus), abbiamo notato una cosa sorprendente: i recettori EGFR rallentavano significativamente! La loro velocità di diffusione diminuiva di circa la metà. E, di nuovo, questo effetto spariva se trattavamo le cellule con sialidasi. Non solo: la frazione di recettori “immobili” o molto lenti aumentava proprio vicino ai virus.
Questo suggerisce che il virus non si limita ad “acchiappare” i recettori EGFR che passano di lì, ma li “trattiene”, forse facilitando la formazione di complessi più stabili necessari per inviare segnali all’interno della cellula. Inoltre, osservando queste zone nel tempo, abbiamo visto che i cluster di EGFR che si formavano erano dinamici: apparivano e scomparivano, indicando un continuo scambio di recettori tra questa zona di contatto e il resto della membrana. L’interfaccia virus-cellula non è una piattaforma statica, ma un’area di intensa e dinamica attività molecolare!
Il Segnale di Ingresso: Reclutare la Macchina dell’Endocitosi
Ok, il virus ha stabilito un contatto, ha rallentato i recettori EGFR… e poi? Il passo successivo è l’ingresso nella cellula tramite endocitosi. Sapevamo che l’IAV usa spesso la via della CME. Una proteina adattatrice chiave in questo processo, per l’IAV, si pensava fosse l’epsina-1. Abbiamo quindi usato cellule che esprimevano epsina-1 fluorescente (epsin1-GFP) e le abbiamo fatte crescere sui nostri virus immobilizzati. Come previsto, abbiamo visto l’epsina-1 accumularsi in corrispondenza di circa il 38% dei virus, un valore simile a quanto riportato in precedenza. Fin qui, tutto tornava.
Ma c’è un’altra proteina adattatrice fondamentale per la CME “classica”, chiamata AP2 (Adaptor Protein 2). Studi precedenti avevano suggerito che l’AP2 *non* fosse coinvolta nell’ingresso dell’IAV. Qui arriva la sorpresa: usando cellule che esprimevano sia AP2 (marcata in rosso, AP2-RFP) sia la clatrina stessa (marcata in verde, ClC-GFP), abbiamo osservato che entrambe le proteine si accumulavano in corrispondenza di oltre il 60% dei virus immobilizzati! Quindi, contrariamente a quanto si pensava, anche AP2 viene reclutata dal virus.

Non solo: monitorando queste cellule vive nel tempo, abbiamo visto che i segnali fluorescenti di clatrina e AP2 fluttuavano. Non era un accumulo statico, ma un continuo “andirivieni”, con picchi di segnale che si ripetevano in media ogni minuto circa (62 secondi per la clatrina, 74 per AP2). Questo indica un’interazione ricorrente tra il virus immobilizzato e la macchina cellulare della CME. È come se la cellula “provasse” ripetutamente a internalizzare il virus, formando e disfacendo le strutture di endocitosi nel sito di legame. Questo dimostra che i nostri virus immobilizzati sono ancora capaci di indurre una risposta cellulare attiva e dinamica.
La Risposta dello Scheletro Cellulare: L’Actina si Riorganizza
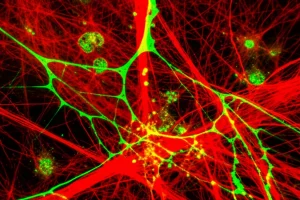
Infine, ci siamo chiesti se anche lo scheletro della cellula, in particolare la rete di filamenti di actina (F-actina) che si trova subito sotto la membrana plasmatica, rispondesse alla presenza del virus. L’actina gioca un ruolo in molti processi cellulari, inclusa l’endocitosi, dove si pensa aiuti a deformare la membrana. Studi precedenti avevano dato risultati contrastanti sul ruolo dell’actina nell’ingresso dell’IAV, ma ricerche più recenti suggerivano una rapida riorganizzazione delle proteine regolatrici dell’actina dopo il contatto con il virus.
Abbiamo quindi marcato l’F-actina nelle nostre cellule (con falloidina fluorescente) e le abbiamo osservate in TIRF. Abbiamo notato che circa il 15% dei virus mostrava un accumulo distinto di F-actina nelle vicinanze. Per vedere meglio questa struttura, siamo passati a un’altra tecnica di super-risoluzione, la STORM (Stochastic Optical Reconstruction Microscopy). Le immagini STORM ci hanno rivelato una rete complessa di filamenti di actina e, vicino ai virus, abbiamo identificato tre tipi principali di organizzazioni nanoscopiche:
- Un accumulo locale, una sorta di “macchia” (blob).
- Un’area priva di segnale, un “buco” (hole).
- Strutture a forma di anello o arco (ring/arc).
Queste strutture ci ricordavano quelle osservate da altri durante la CME. Abbiamo ipotizzato che potessero essere istantanee diverse dello stesso processo dinamico. Per verificarlo, siamo tornati all’imaging di cellule vive, questa volta usando cellule che esprimevano LifeAct-GFP (una piccola proteina che si lega all’F-actina senza disturbarla troppo). E infatti, osservando il segnale di LifeAct-GFP vicino ai singoli virus nel tempo, abbiamo visto proprio questo: fluttuazioni dinamiche e transizioni tra strutture simili a quelle viste in STORM (buchi, anelli, macchie)! L’accumulo locale di actina era spesso di breve durata, il che spiega perché lo vedevamo solo nel 15% dei casi nelle cellule fissate.

Cosa Abbiamo Imparato (e Cosa Ci Riserva il Futuro)
Quindi, mettendo insieme tutti i pezzi, il nostro lavoro mostra che un singolo virus influenzale, una volta attaccato alla cellula, orchestra una complessa serie di eventi nanoscopici proprio nell’interfaccia virus-cellula:
- Rallenta i recettori EGFR in modo dipendente dall’acido sialico, favorendone l’accumulo dinamico.
- Induce il reclutamento della macchina dell’endocitosi mediata da clatrina, coinvolgendo non solo l’epsina-1 ma, sorprendentemente, anche l’AP2, con dinamiche ricorrenti.
- Provoca una riorganizzazione locale e dinamica del citoscheletro di actina, con la formazione transitoria di strutture specifiche (buchi, anelli, macchie).
Il nostro metodo di immobilizzazione dei virus si è rivelato uno strumento potente per “dissezionare” questi primissimi istanti dell’infezione con un dettaglio senza precedenti. Ci permette di studiare l’interazione tra la cellula e più virus contemporaneamente e può essere adattato a diverse tecniche di microscopia avanzata.
Questa comprensione più profonda delle strategie che i virus usano per entrare nelle nostre cellule è fondamentale. Ogni passaggio che scopriamo potrebbe diventare un potenziale bersaglio per nuovi farmaci antivirali. Inoltre, questa tecnica non è limitata all’influenza: potrebbe essere applicata per studiare l’ingresso di molti altri virus, batteri o persino nanoparticelle sintetiche, aprendo nuove strade nella ricerca sulle infezioni e sulla biologia cellulare a livello della singola molecola. È davvero un momento entusiasmante per chi, come noi, ama spiare i segreti più intimi delle cellule!
Fonte: Springer Nature