Vibrio Cholerae: Come il Batterio del Colera Schiva i Nostri Proiettili Magici!
Ciao a tutti, appassionati di scienza e misteri del microcosmo! Oggi voglio portarvi con me in un viaggio affascinante nel mondo dei batteri, in particolare di uno piuttosto famoso (e famigerato): il Vibrio cholerae, l’agente causale del colera. Ma non parleremo della malattia in sé, bensì di una scoperta incredibile che riguarda le sue strategie di sopravvivenza. Immaginate i batteri come piccoli guerrieri che devono difendersi costantemente dagli attacchi. Uno degli attacchi più comuni proviene dai cosiddetti peptidi antimicrobici (AMP).
Cosa sono questi AMP?
Pensate agli AMP come a delle minuscole “pallottole magiche” prodotte sia dalle nostre cellule (per combattere le infezioni) sia da altri batteri (per farsi spazio e competere per le risorse). Sono piccole molecole cariche positivamente che adorano attaccarsi alla superficie dei batteri, che è carica negativamente. Questo incontro spesso provoca buchi nella membrana batterica, facendo fuoriuscire il contenuto interno e… addio batterio! Alcuni AMP, come la Polimixina B (PmB), sono così potenti che li usiamo come farmaci di ultima istanza contro infezioni batteriche super resistenti. La PmB è prodotta da un altro batterio, il Paenibacillus polymyxa, ed è un po’ un’arma a doppio taglio: utile in medicina, ma il suo uso (anche negli allevamenti) può favorire lo sviluppo di resistenze.
Il Nostro Indagato Speciale: Vibrio cholerae
Il Vibrio cholerae vive principalmente in ambienti acquatici, ma quando entra nel nostro corpo attraverso acqua o cibo contaminati, può scatenare il colera. Questo batterio non è sprovveduto; sa difendersi. Ad esempio, alcuni ceppi (come gli El Tor, responsabili della pandemia attuale) sanno come modificare la loro superficie esterna (il lipopolisaccaride o LPS) per renderla meno “attraente” per gli AMP come la PmB. Questo meccanismo principale di resistenza coinvolge un sistema chiamato almEFG, la cui attivazione è regolata da un altro sistema a due componenti, CarRS (noto anche come VprAB), che “sente” la presenza di AMP nel periplasma (lo spazio tra le due membrane batteriche). Ma non è tutto qui… ci sono anche le pompe di efflusso, meccanismi che letteralmente “sputano fuori” le sostanze tossiche, come la pompa VexAB.
La Scoperta Inaspettata: OmpV Entra in Scena
Ed è qui che la nostra storia si fa interessante. Studiando come il V. cholerae (in particolare il ceppo El Tor A1552) reagisce alla PmB a livello di espressione genica (cioè quali geni vengono “accesi” o “spenti”), abbiamo notato qualcosa di strano. Un gene, chiamato ompV, che codifica per una proteina della membrana esterna fino ad allora poco caratterizzata, veniva attivato massicciamente in presenza di PmB! Questa proteina, OmpV, può diventare addirittura una delle più abbondanti sulla superficie del batterio in certe condizioni. Si sapeva che poteva avere un ruolo nella patogenesi (il nostro sistema immunitario produce anticorpi contro di essa durante l’infezione), ma il suo legame con la resistenza agli AMP era tutto da scoprire.

Un Operone Specializzato per la Difesa
Guardando più da vicino il genoma del V. cholerae, abbiamo fatto un’altra scoperta chiave: il gene ompV non è solo! Si trova raggruppato insieme ad altri tre geni in un’unica unità funzionale chiamata operone. E chi sono i suoi compagni di viaggio? Proprio il sistema a due componenti carRS di cui parlavamo prima, e un altro gene chiamato virK, la cui funzione in V. cholerae era sconosciuta, ma che in altri batteri è legato alla virulenza e alla resistenza agli AMP. Questo quartetto, l’operone carRS-ompV-virK, è incredibilmente conservato in tantissimi ceppi di V. cholerae, suggerendo che sia molto importante. Abbiamo confermato con esperimenti di RT-PCR che questi geni vengono trascritti tutti insieme sullo stesso “nastro” di mRNA e che l’intero operone viene attivato dalla presenza di PmB.
OmpV e VirK: Guardie del Corpo Batteriche
A questo punto, la domanda era ovvia: se ompV e virK vengono attivati insieme a carRS (già noto per la resistenza), avranno anche loro un ruolo difensivo? Per scoprirlo, abbiamo creato dei mutanti, batteri a cui avevamo “tolto” il gene ompV o il gene virK (o entrambi). Ebbene sì! I batteri senza ompV (A1552ΔompV) e quelli senza virK (A1552ΔvirK) erano significativamente più sensibili sia alla Polimixina B che a un altro AMP umano, LL-37. Reinserendo i geni mancanti (complementazione), la resistenza veniva ripristinata, confermando il loro ruolo. Curiosamente, il mutante doppio (senza entrambi i geni) non era più sensibile del mutante senza solo ompV, suggerendo che OmpV potrebbe avere un ruolo predominante o che agiscano sulla stessa via.
Ma Come Funziona OmpV? Non è Solo Questione di Membrana o “Scudi”
Ok, OmpV aiuta nella resistenza. Ma come? La prima ipotesi poteva essere che, essendo una proteina abbondante della membrana esterna, la sua assenza destabilizzasse la membrana, rendendola più vulnerabile. Abbiamo testato questa idea usando sonde fluorescenti (NPN e PI) che si illuminano se la membrana è danneggiata. Risultato? Nessuna differenza significativa tra i batteri normali e quelli senza OmpV quando esposti a PmB. La membrana non sembrava più “fragile” senza OmpV. Quindi, la sensibilità non era dovuta a una destabilizzazione generale.
Un’altra idea riguardava le vescicole di membrana (MV). I batteri rilasciano queste piccole “bolle” dalla loro superficie, che possono agire come esche, catturando gli AMP prima che raggiungano il batterio vero e proprio. Avevamo visto in studi precedenti che OmpV è abbondante nelle MV rilasciate in presenza di AMP. Forse senza OmpV, i batteri producevano meno MV o MV meno “appiccicose” per la PmB? Abbiamo misurato la produzione di MV e testato la loro capacità protettiva: nessuna differenza rilevante tra i batteri normali e quelli senza OmpV. Quindi, anche se OmpV è nelle MV, il suo ruolo cruciale nella resistenza sembra essere un altro.

Il Colpo di Scena: OmpV Controlla un’Altra Arma!
Se OmpV non agisce destabilizzando la membrana o tramite le MV, cosa fa? Siamo tornati ai dati di espressione genica, ma questa volta confrontando i batteri normali e quelli senza ompV, entrambi esposti a PmB. Ricordate la pompa di efflusso VexAB, un altro meccanismo di resistenza? Nei batteri normali, la PmB ne aumentava l’espressione, come previsto. Ma nei batteri senza ompV… sorpresa! L’espressione di vexB (la componente principale della pompa) non aumentava in risposta alla PmB. Ecco la chiave! Sembra che OmpV sia necessario per “dare il segnale” di attivare la pompa VexAB quando arriva la minaccia PmB. Questo è importantissimo: OmpV non è solo una difesa passiva, ma partecipa attivamente alla regolazione di altri sistemi di resistenza! L’attivazione di altri geni legati alla resistenza, come almG (controllato da CarRS) e rpoE (un fattore di risposta allo stress), avveniva ancora (anche se l’attivazione di carS era un po’ ridotta senza ompV), indicando che OmpV controlla specificamente (o almeno in modo predominante) la via che porta a VexAB.
La Struttura di OmpV: Un Sensore con una “Tasca Segreta”?
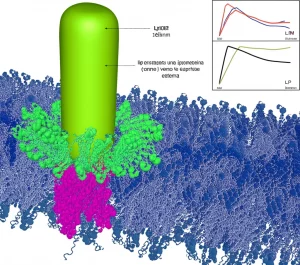
Ma come fa una proteina sulla superficie esterna a controllare l’espressione di geni all’interno? Qui entra in gioco la struttura tridimensionale della proteina OmpV, che abbiamo predetto usando potenti strumenti computazionali come AlphaFold2. La previsione è affascinante: OmpV forma una struttura a barile beta (comune per le proteine della membrana esterna), ma con una particolarità unica. Due delle “doghe” del barile (i filamenti beta 5 e 6) sono più corte e lasciano un’apertura laterale! Questa apertura dà accesso a una tasca all’interno del lume del barile, una tasca che è isolata sia dall’esterno che dal periplasma. E indovinate un po’? Questa tasca è fortemente elettronegativa (carica negativamente). La Polimixina B è un peptide cationico (carico positivamente). Vedete dove voglio arrivare? Simulazioni di docking (che prevedono come le molecole si legano) suggeriscono che la PmB può infilarsi perfettamente in questa tasca elettronegativa attraverso l’apertura laterale, con un’alta affinità.

L’ipotesi, quindi, è che OmpV funzioni come un sensore. Quando la PmB si integra nella membrana esterna, può scivolare nell’apertura laterale di OmpV e legarsi alla tasca interna. Questo legame potrebbe indurre un cambiamento conformazionale nella proteina OmpV, un po’ come una chiave che gira in una serratura. Questo cambiamento strutturale potrebbe poi innescare una cascata di segnali all’interno della cellula (forse coinvolgendo VirK come mediatore periplasmatico? O interagendo con CarS?), portando infine all’attivazione della pompa di efflusso VexAB.
Un Operone, Molteplici Difese: Una Strategia Sofisticata
Quello che abbiamo scoperto è davvero notevole. Il Vibrio cholerae possiede un operone specializzato, carRS-ompV-virK, che orchestra l’attivazione di molteplici sistemi di resistenza agli AMP in risposta alla minaccia:
- Il sistema CarRS attiva la modifica dell’LPS tramite almEFG (per ridurre l’attrazione degli AMP).
- La proteina OmpV (probabilmente agendo da sensore) è necessaria per attivare la pompa di efflusso VexAB (per espellere gli AMP che entrano).
- Anche VirK contribuisce alla resistenza, anche se il suo ruolo esatto nel meccanismo di segnalazione resta da chiarire.
È la prima volta che viene identificato in V. cholerae un singolo locus genetico (diverso da un semplice sistema a due componenti) che regola sistemi di resistenza così diversi. È una strategia difensiva complessa ed efficiente, conservata in molti ceppi, che aiuta questo batterio a sopravvivere agli attacchi chimici nel suo ambiente e, potenzialmente, nel nostro corpo.
Capire questi meccanismi è fondamentale nella lotta contro la resistenza agli antibiotici. Identificare i sensori come OmpV e i meccanismi di segnalazione potrebbe aprire la strada a nuove strategie terapeutiche per rendere i batteri nuovamente vulnerabili ai nostri farmaci. Il piccolo Vibrio ci ha svelato un altro dei suoi trucchi, e noi siamo qui per studiarli e, si spera, per trovare un modo per contrastarli!
Fonte: Springer