MetQ: La Chiave Nascosta di Streptococcus suis tra Virulenza e Misteriosi Biofilm!
Amici appassionati di scienza, oggi vi porto nel mondo un po’ losco ma affascinante dei batteri, e in particolare di uno che sa il fatto suo quando si tratta di causare problemi: lo Streptococcus suis. Se avete a che fare con suini o siete semplicemente curiosi di come questi microrganismi riescano a essere così… efficaci, mettetevi comodi, perché stiamo per svelare uno dei loro piccoli, sporchi segreti.
Immaginate questo batterio come un piccolo ma agguerrito guerriero. Per sopravvivere e prosperare, soprattutto quando cerca di infettare un ospite (come i nostri amici maiali, o sfortunatamente a volte anche noi umani), ha bisogno di nutrienti. E uno di questi, fondamentale, è la metionina. Sì, proprio quell’amminoacido essenziale di cui sentiamo parlare anche per la nostra dieta. Ecco, S. suis non fa eccezione, ne ha un bisogno vitale.
Ma come fa a procurarsela? Ecco che entra in gioco MetQ!
Nel nostro laboratorio (metaforicamente parlando, visto che vi sto raccontando i risultati di uno studio specifico!), ci siamo chiesti: come fa S. suis a “mangiare” la metionina quando l’ambiente scarseggia? Abbiamo messo sotto la lente d’ingrandimento un sospettato principale: un sistema di trasporto chiamato MetQ. Pensatelo come una specie di “porta di servizio” specializzata, o meglio, una sofisticata pompa molecolare. Più tecnicamente, MetQ è parte di un complesso chiamato trasportatore ABC, un meccanismo super efficiente che i batteri usano per importare sostanze vitali.
Abbiamo scoperto che il gene metQ è parte di un gruppetto di geni (un operone, per i più tecnici) che include anche geni per una transposasi e un’ATPasi. Insieme, formano un classico trasportatore ABC tripartito. La cosa interessante è che questa configurazione è piuttosto comune in diverse specie di streptococchi, sia quelli che colpiscono gli animali sia quelli che infettano l’uomo. Una sorta di “design” di successo nel mondo batterico.
MetQ: Carta d’identità e localizzazione
Per prima cosa, volevamo essere sicuri che MetQ fosse davvero lui, il “portiere” della metionina. Utilizzando tecniche come il Western blotting e la citometria a flusso (con un antisiero specifico che abbiamo sviluppato, una specie di “segugio” molecolare per MetQ), abbiamo visto che S. suis (in particolare il ceppo di riferimento P1/7) produce MetQ quando si trova in condizioni di carenza di metionina. E indovinate un po’ dove si trova? Proprio sulla superficie della cellula batterica, pronto ad acchiappare la metionina di passaggio. Logico, no?
Per confermare ulteriormente il suo ruolo, abbiamo fatto crescere il batterio in terreni di coltura definiti chimicamente, dove potevamo controllare con precisione la quantità di metionina. Abbiamo creato un mutante di S. suis a cui avevamo “spento” il gene metQ (chiamiamolo P1/7∆metQ). Ebbene, questo mutante faceva molta fatica a crescere quando la metionina era scarsa. Se invece il terreno era ricco di nutrienti, cresceva normalmente. Questo ci ha detto chiaramente che MetQ è il principale responsabile del trasporto della metionina in condizioni di “carestia”.
Non contenti, abbiamo voluto misurare quanto “forte” MetQ lega la metionina. Con una tecnica chiamata Calorimetria di Titolazione Isoterma (ITC), abbiamo visto che MetQ si lega alla L-metionina con una costante di dissociazione (KD) di 7.1 µM. Tradotto: ha una buona affinità per il suo “cibo”.
Il ruolo di MetQ nella virulenza: un fattore chiave
Ora, la parte che forse interessa di più: MetQ c’entra qualcosa con la capacità del batterio di fare danni, cioè con la sua virulenza? Per scoprirlo, abbiamo usato un modello di infezione murino (poveri topolini, ma sono fondamentali per la ricerca!). Abbiamo visto che il nostro mutante P1/7∆metQ aveva una capacità ridotta di diffondersi agli organi interni rispetto al batterio “normale” (wild type). Un chiaro segnale che MetQ è importante per l’infezione.
Ma perché? Un’ipotesi è legata alla sopravvivenza all’interno delle cellule del sistema immunitario, i macrofagi, che sono un po’ gli “spazzini” del nostro corpo. Il mutante P1/7∆metQ mostrava una ridotta sopravvivenza intracellulare nei macrofagi murini. Inoltre, era più sensibile allo stress ossidativo. I macrofagi, infatti, producono sostanze ossidanti per uccidere i batteri invasori. Se il batterio non riesce a procurarsi abbastanza metionina (che ha anche ruoli antiossidanti indiretti e serve per sintetizzare proteine vitali per la difesa), diventa un bersaglio più facile.
Quindi, MetQ non è solo un “fattorino” della metionina, ma un elemento cruciale per la nutrizione batterica, per sfuggire alle difese immunitarie e, in definitiva, per la patogenicità durante l’infezione. Senza un adeguato apporto di metionina, garantito da MetQ, S. suis è meno equipaggiato per la battaglia.
La sorpresa: MetQ e la formazione di biofilm
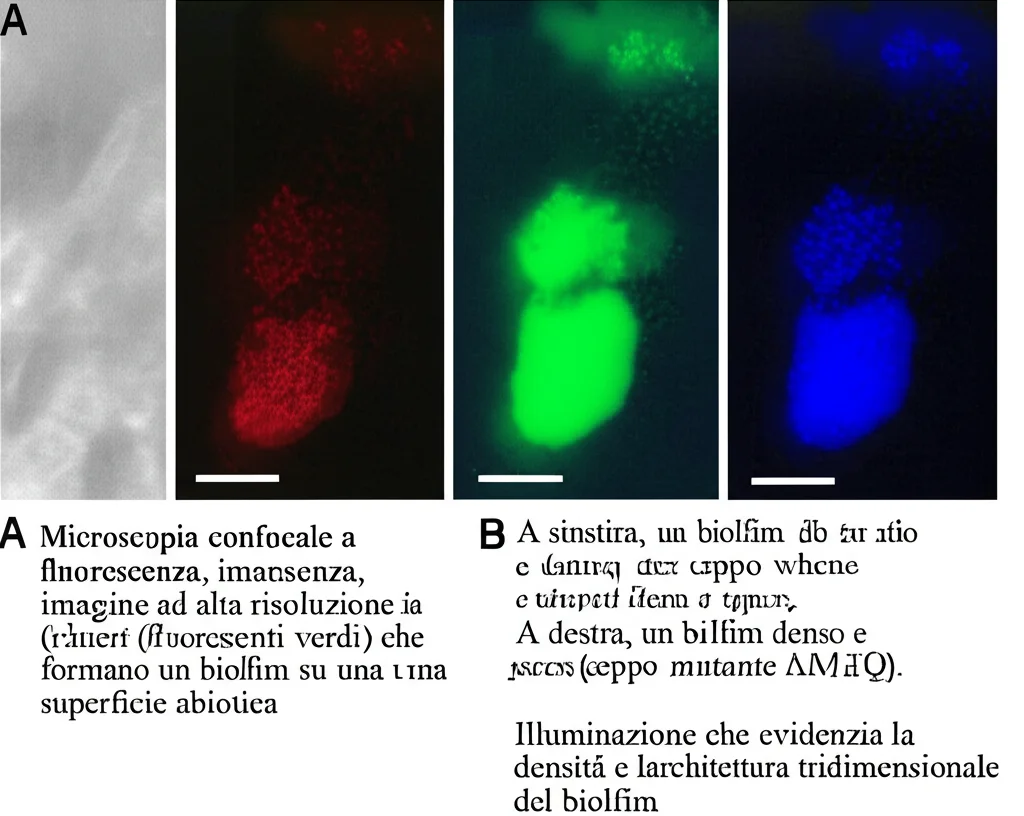
E qui arriva il colpo di scena! Ci aspettavamo che un batterio “azzoppato” dalla mancanza di MetQ fosse meno bravo a fare un po’ tutto. Invece, abbiamo osservato una cosa curiosa: il mutante P1/7∆metQ mostrava una formazione di biofilm potenziata rispetto al wild type. I biofilm, per chi non lo sapesse, sono quelle comunità strutturate di batteri adese a una superficie, un po’ come delle “cittadelle” batteriche che li proteggono da stress ambientali e antibiotici. È un risultato controintuitivo: un batterio meno virulento in un certo senso, ma più bravo a “costruire case”.
Abbiamo esaminato l’architettura di questi biofilm con microscopia confocale. I biofilm formati da P1/7 (quello con MetQ funzionante) mostravano cluster batterici dispersi con spazi significativi tra loro. Al contrario, i biofilm prodotti dal mutante P1/7∆metQ presentavano grandi cluster batterici interagenti che coprivano una superficie maggiore. Analisi bioinformatiche hanno confermato che i biofilm del mutante erano significativamente più spessi, avevano un coefficiente di rugosità inferiore e occupavano un’area maggiore.
Questo suggerisce che MetQ, o meglio la disponibilità di metionina che esso regola, gioca un ruolo complesso e forse inaspettato anche nella modulazione della struttura e della dinamica del biofilm. Potrebbe essere che, in condizioni di stress nutrizionale (come la difficoltà a importare metionina), il batterio “decida” di rintanarsi e formare biofilm più robusti come strategia di sopravvivenza alternativa?
Cosa ci dice tutto questo?
Beh, che la vita dei batteri è incredibilmente complessa e finemente regolata. La metionina non è solo un mattoncino per le proteine, ma la sua acquisizione influenza profondamente il metabolismo, la capacità di causare malattie e persino il comportamento sociale dei batteri, come la formazione di biofilm.
Lo studio del gene metQ (noto anche come SSU1577 nel ceppo di riferimento) ci ha aperto una finestra importante. Abbiamo visto che la sua organizzazione genica in S. suis (metQ seguito da dapE, metN, e metI) è un po’ diversa dal “canone” visto in altri batteri, dove metQ è spesso alla fine dell’operone. Questa particolarità potrebbe indicare riarrangiamenti genetici specifici in Streptococcus. La presenza di dapE (coinvolto nella sintesi della lisina ma con possibile attività peptidolitica) nell’operone della metionina è intrigante: potrebbe aiutare a “liberare” metionina da piccoli peptidi?
La ridotta virulenza del mutante ∆metQ nei topi conferma quanto già sospettato da studi precedenti su altri ceppi e in modelli suini. La metionina è scarsa nei fluidi corporei dei mammiferi (1-7 µg/mL), quindi un trasportatore efficiente come MetQ è vitale. Senza di esso, S. suis cresce meno, forse esprime meno fattori di virulenza come la capsula (che lo protegge dalla fagocitosi), e soccombe più facilmente allo stress ossidativo inflitto dalle cellule immunitarie.
Riguardo al biofilm, è la prima volta che si dimostra un ruolo di MetQ in questo processo per S. suis, e in modo così particolare (la sua assenza lo aumenta!). In altri batteri, come Neisseria gonorrhoeae, MetQ sembra agire come un’adesina, quindi la sua assenza ridurrebbe il biofilm. Qui è il contrario. Una possibile spiegazione potrebbe risiedere nei livelli intracellulari di cAMP. È noto che la restrizione di metionina può aumentare il cAMP, un secondo messaggero che in molti patogeni promuove la formazione di biofilm stimolando la produzione di adesine come i pili di tipo IV (presenti anche negli streptococchi). È una speculazione, certo, ma affascinante!
In conclusione, le nostre scoperte (o meglio, quelle dello studio che vi ho raccontato!) indicano che MetQ è essenziale per l’assorbimento della metionina in condizioni di carenza, il che è critico per la nutrizione batterica, l’evasione immunitaria e la patogenicità durante l’infezione. E, come ciliegina sulla torta, sembra essere un modulatore inaspettato della formazione di biofilm. Capire questi meccanismi non è solo un esercizio accademico: potrebbe aprire la strada a nuove strategie per combattere questo patogeno, magari prendendo di mira proprio il suo sistema di approvvigionamento di metionina. La ricerca continua, e chissà quali altri segreti ci svelerà Streptococcus suis!
Fonte: Springer





