Ciliati: Come la Danza dell’Acqua Scolpisce i Predatori Microscopici
Ciao a tutti! Oggi voglio portarvi in un mondo invisibile a occhio nudo, ma brulicante di vita e di strategie ingegnose: il mondo dei ciliati. Immaginate organismi unicellulari, a volte mille volte più grandi di un batterio, che nuotano o si ancorano in una goccia d’acqua. Sembrano semplici, vero? Eppure, nascondono segreti affascinanti su come la vita stessa si è evoluta per… mangiare!
La Sfida di Nutrirsi nel Microcosmo
Pensateci: per le primissime forme di vita, come batteri e archei, nutrirsi significava assorbire molecole disciolte nell’ambiente attraverso la membrana cellulare. Un processo chiamato diffusione, efficace solo su piccolissime scale. Questo limitava drasticamente le dimensioni che potevano raggiungere. Poi, è successa una cosa rivoluzionaria: la fagotrofia. È la capacità di inghiottire particelle di cibo intere, altri organismi inclusi! Questo ha aperto le porte all’evoluzione degli eucarioti unicellulari, cellule più complesse con un nucleo definito, e alla fine, alla vita multicellulare come la conosciamo.
Ma inghiottire particelle pone una sfida meccanica non da poco: come coordinare il trasferimento del cibo dall’ambiente esterno all’interno della cellula? È qui che entrano in gioco i nostri amici ciliati.
I Ciliati: Artisti del Movimento Acquatico
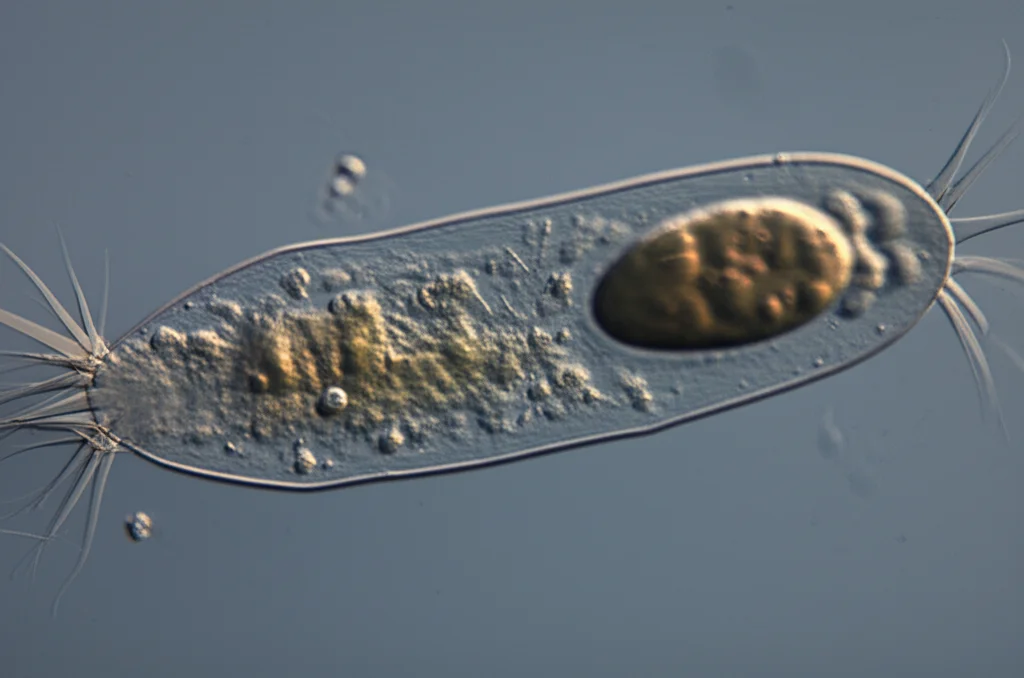
I ciliati sono protisti acquatici caratterizzati da minuscole appendici simili a peli, le cilia. Queste cilia non sono ferme, anzi! Si muovono in modo coordinato come migliaia di piccoli remi, permettendo alla cellula di nuotare o, se è ancorata (sessile), di creare correnti d’acqua per attirare il cibo. Queste correnti convogliano le particelle verso una sorta di “bocca” specializzata, il citostoma, da dove il cibo viene ingerito e poi processato internamente.
È incredibile pensare che gli stessi materiali di base – microtubuli e motori molecolari come la dineina – che formano lo “scheletro” interno della cellula e permettono il trasporto al suo interno, compongono anche le cilia che gestiscono il flusso esterno. Questo suggerisce una profonda co-evoluzione tra il trasporto interno ed esterno.
Ma osservando i ciliati, notiamo una cosa curiosa: quelli che nuotano liberamente (come il famoso Paramecium) hanno spesso cilia distribuite su gran parte della superficie, mentre quelli sessili (come l’elegante Stentor a forma di tromba) tendono ad avere le cilia concentrate in bande specifiche attorno alla zona della bocca. Perché questa differenza? La disposizione ottimale delle cilia dipende forse dallo stile di vita?
La Fisica ci Svela il Segreto: Diffusione vs. Flusso Attivo
Per capire, dobbiamo immergerci un po’ nella fisica dei fluidi su microscala. In questo mondo dominato dalla viscosità (il numero di Reynolds è bassissimo, l’inerzia conta poco), il trasporto dei nutrienti può avvenire per diffusione o tramite movimento attivo del fluido.
Per i batteri piccolissimi, la diffusione è sovrana. Aumentare il flusso d’acqua intorno a loro non migliora significativamente l’assorbimento di nutrienti disciolti. Anzi, nuotare ha un costo energetico. La strategia migliore per loro è avere recettori distribuiti uniformemente su tutta la superficie.
Ma per cellule più grandi, come i ciliati, le cose cambiano. Il numero di Péclet (Pe) ci dice quanto è importante il trasporto dovuto al flusso rispetto alla diffusione. Più grande è la cellula, più vantaggioso diventa creare attivamente correnti d’acqua. E qui sta il bello: il costo energetico per migliorare l’assorbimento di nutrienti tramite il flusso *diminuisce drasticamente* all’aumentare delle dimensioni della cellula (scala come 1/a⁴, dove ‘a’ è la dimensione!). Per un ciliato di 100 micron, rispetto a un batterio di 1 micron, il costo relativo si riduce di 10.000 volte!

Ecco perché per i ciliati vale la pena “investire” energia nel mantenere attive le cilia per creare correnti alimentari. Allo stesso tempo, affidarsi solo alla diffusione per trasportare le sostanze all’interno di una cellula grande diventerebbe troppo lento. Ed ecco che entra in gioco il trasporto attivo interno basato sul citoscheletro. Fisica ed evoluzione vanno a braccetto!
Modellare l’Efficienza: Trovare il Design Perfetto
Ma qual è la disposizione ottimale di bocca e cilia per massimizzare l’arrivo del cibo? Per rispondere, abbiamo usato modelli matematici. Immaginate la cellula come una sfera immersa in un fluido viscoso. Abbiamo assegnato una frazione della superficie (Af) alla “bocca” (zona assorbente) e una frazione adiacente (Ac) alle cilia attive, mantenendo costante l’energia spesa per muovere le cilia. Abbiamo poi calcolato due misure di performance: il flusso alimentare (Q), cioè quanto fluido viene diretto verso la bocca, e il numero di Sherwood (Sh), che confronta l’assorbimento di nutrienti con quello che avverrebbe per sola diffusione.
Risultati Sorprendenti: Strategie Diverse per Stili di Vita Diversi
I risultati sono stati illuminanti!
- Per i ciliati sessili (attaccati): L’efficienza massima (sia per Q che per Sh) si ottiene quasi sempre concentrando le cilia in una banda stretta proprio attorno alla bocca. Questo non solo massimizza l’arrivo del cibo, ma minimizza anche la forza necessaria per rimanere ancorati al substrato. Una doppia vittoria!
- Per i ciliati motili (nuotatori): La situazione è più complessa. Se la bocca è relativamente piccola (Af sotto il 20% circa), anche per loro le bande di cilia attorno alla bocca sono ottimali. Ma se la bocca occupa una frazione maggiore della superficie, l’ottimo “biforca”: diventa più vantaggioso coprire di cilia tutta la superficie rimanente non occupata dalla bocca. Questo design massimizza sia l’assorbimento di nutrienti sia la velocità di nuoto.
Abbiamo anche verificato che mettere le cilia proprio *adiacenti* alla bocca è sempre meglio che lasciare uno spazio inattivo tra le due zone. E un’altra scoperta interessante: in entrambi i casi, l’area ottimale della bocca (Af) è sempre inferiore al 50% della superficie totale. Avere una “bocca” dedicata è meglio che cercare di assorbire ovunque quando si usa il flusso attivo.

Il Mondo Reale Conferma il Modello (con una Sorpresa)
Emozionati da questi risultati teorici, siamo andati a vedere cosa succede in natura. Abbiamo analizzato i dati morfologici di 31 generi diversi di ciliati, sia sessili che motili. Abbiamo stimato le loro frazioni di superficie dedicate alla bocca (Af) e alle cilia (Ac) e le abbiamo mappate sul nostro “spazio dei design” teorico.
I risultati sono stati straordinari!
- I ciliati sessili reali (come Stentor, Vorticella) si raggruppano proprio nella regione del nostro modello che prevede bande ciliari localizzate come strategia ottimale.
- I ciliati motili reali si dividono in due gruppi, proprio come previsto dalla biforcazione del nostro modello! Alcuni (come Didinium) hanno bande ciliari localizzate, simili ai sessili. Altri (come Paramecium, Tetrahymena) hanno cilia che coprono quasi tutta la superficie non orale, seguendo l’altra strategia ottimale che bilancia nuoto e alimentazione.
La fisica dei fluidi sembra davvero scolpire la forma e la funzione di questi organismi!
Oltre l’Ottimizzazione: Il Principio del “Sufficientemente Buono”?
C’è però un ultimo tassello intrigante. Quando abbiamo calcolato i tassi di assorbimento di nutrienti (Q e Sh) previsti per i design dei ciliati reali, abbiamo scoperto che tutti, indipendentemente dalla loro strategia (bande localizzate o copertura totale), si trovano in una regione del nostro spazio dei design dove l’assorbimento è già molto alto: tipicamente più di 10.000 volte il volume corporeo all’ora per Q, e più del doppio rispetto alla sola diffusione per Sh.
Questo è affascinante. Sembra che, una volta superata una certa soglia di efficienza (diciamo, raddoppiare l’assorbimento rispetto alla diffusione), un ulteriore aumento dell’efficienza nel catturare cibo dall’esterno non sia più la forza selettiva dominante. Perché? Le ipotesi sono diverse:
- Forse il vero limite non è catturare il cibo, ma la capacità interna della cellula di processarlo.
- Forse i costi energetici per costruire e mantenere *più* strutture ciliari diventano proibitivi.
- Forse altri processi evolutivi, come la deriva genetica (importante in organismi con piccole popolazioni efficaci come i protisti), hanno giocato un ruolo.
- O forse, semplicemente, un aumento di due volte nell’assorbimento di nutrienti è “sufficientemente buono” per la sopravvivenza e la competizione, e non c’è un forte vantaggio selettivo nell’ottimizzare ulteriormente questo aspetto.
 2. Stile da pubblicazione scientifica, chiaro e informativo.” />
2. Stile da pubblicazione scientifica, chiaro e informativo.” />
Conclusioni: Una Danza tra Fisica ed Evoluzione
Quello che abbiamo scoperto è una mappa quantitativa che lega il modo in cui i ciliati si nutrono alla loro “architettura” superficiale. Il passaggio da piccole cellule dipendenti dalla diffusione a eucarioti più grandi capaci di fagotrofia ha richiesto nuove strategie per acquisire nutrienti. Abbiamo visto come la fisica renda energeticamente vantaggioso per le cellule più grandi usare le cilia per creare flussi alimentari.
La diversità nelle disposizioni delle cilia non è casuale, ma è plasmata dalla fisica dei fluidi in relazione allo stile di vita: bande concentrate per i sessili (ottimo per mangiare restando fermi), e strategie biforcate per i motili (bilanciando cibo e movimento). Il fatto che tutti raggiungano livelli di assorbimento simili, pur potendo teoricamente fare di più (specialmente alcuni motili), suggerisce che l’evoluzione è un complesso gioco di equilibri, dove l’ottimizzazione di un singolo aspetto può non essere l’unico obiettivo.
Capire queste dinamiche non è solo affascinante di per sé, ma ci aiuta a comprendere l’ecologia dei protisti, il funzionamento delle catene alimentari acquatiche e persino le basi della cooperazione tra cellule, un passo fondamentale verso la multicellularità. La prossima volta che penserete a una semplice goccia d’acqua, ricordatevi della complessa e meravigliosa danza della fisica e dell’evoluzione che avviene al suo interno!
Fonte: Springer